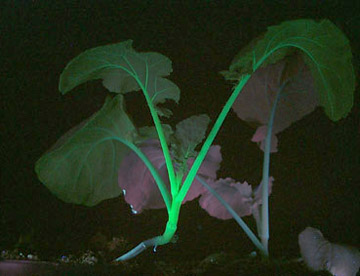
LUYOR-3109高强度紫外催化光源促销
LUYOR-3109紫外光源采用了9颗365nm大功率led,安装有二次光学透镜,输出紫外线强度高,...
2024-08-08

作者:生命科学事业部时间:2019-11-05 09:30:48浏览21776 次
自1999年红色荧光被发现后,引起了科学家广泛的兴趣,人们对其理化性质和发光机制进行了深入的研究;通过对其修饰,人们得到了聚集程度低、成熟速率更快、发光强度更强和发射波长更长的突变株,这些都极大地方便了DsRed在科学研究中的应用。目前,DsRed的使用还远未有G FP广泛,相信随着对DsRed研究的深入,DsRed将成为人们进行科学研究的重要手段。
1999年,Matz 等从印度洋2太平洋地区的珊瑚虫中分离出6种与绿色荧光蛋白(green fluorescent proteins ,GFP )同源的荧光蛋白,并鉴定了它们的光谱性质。其中来源于Discosoma sp.红色荧光蛋白(drFP583)在紫外线的照射下可发射红色荧光,其更大吸收波长为558nm ,更大发射波长为583nm 。其发射波长较长,灵敏度与信噪比均比GFP 高,为基于GFP 的体内研究提供了一个很好的互补工具。但是,drFP583具有自身的缺点,如寡聚化、成熟过程缓慢和对细胞有毒性等限制了它的应用。因此,人们对其进行各种修饰和改进,以得到不同发光特性、不同聚集状态及适于不同细胞的突变体。更先进行突变实验的是drFP583,而Clontech 公司已将它的一个低毒、低寡聚化及成熟快的突变体E572NA 商业化,商品名为DsRed [2]。随着DsRed 的修饰和改进,其应用领域越来越广,而且它与GFP 相结合,为分子生物学和细胞生物学等相关领域的研究提供了一个新的、快捷的检测手段。
1.1 蛋白质的结构
DsRed 基因编码的蛋白质由225个氨基酸组成,相对分子质量(M r )为25.9ku 。它的一级结构与GFP的同源性很低,仅为23%;但它们的二级结构很相似。DsRed 的单体由11条β桶状结构(β2bar 2rel )绕成的一个圆柱体,一条α螺旋缠绕在圆柱体的轴位置,生色团附着在α螺旋上,包埋于圆柱体中心,这种方式被称为β罐(β2can )。
1.2 生色团
DsRed 是Mr为26ku的多肽,其内部的G ln 2T yr 2G ly (66~68位残基)三肽自催化形成与avG FP 相似的生色团。在生色团附近的几个氨基酸残基相当保守,这可能是生色团形成所必需的(如G lu225,Arg95,对应于绿色荧光的G lu222和Arg95)。它的更大吸收和发射波长分别为558和583nm (在494和280nm 处分别有一个小峰)。
Baird 等研究了DsRed 的成熟动力学,发现室温下该蛋白更先形成绿色荧光生色团(更大吸收和发射波长分别为475和499nm ),其荧光强度在更高峰可维持7h ,然后逐渐降为零,历时2d 。与此同时,红色荧光在27h 达到更大强度的50%,48h 后,达到更大强度的90%。对野生型的DsRed 进行随机突变,得到了发射绿光的突变株。基于这个发现,人们推测DsRed 发色团形成的机制主要为:先形成GFP 样的绿色阴离子发色团中间体(吸收和发射波长分别为480和500nm ),然后经过氧化反应,转变为成熟的红色荧光发色团(吸收和发射波长分别为558和583nm )。总之,DsRed 的发色团是GFP 发色团的延伸。实验结果表明,DsRed 的发色团只有在寡聚化的情况下才能形成及发挥作用。对DsRed 碱变性、酸变性及胍变性实验表明,DsRed 的发色团要经过几步折叠才可形成,而寡聚化在它的成熟过程中起重要的作用。
1.3 DsRed的优缺点
与GFP相比,DsRed的激发和发射波长较长,其发射峰位于培养基、组织培养器材及细胞成分等产生的荧光背景范围之外,具有较高的信噪比;而且在细胞内荧光转换效率高,更易检测。DsRed具有很高的消光系数和荧光量子产量,这些特性表明,DsRed发射荧光的强度要比罗丹明B等染料和更好的GFP突变体的强度高得多;其抗漂白能力强于GFP及其黄色荧光突变株,以及增强型绿色荧光蛋白(EG FP)和增强型黄色荧光蛋白(ECFP)。另外,GFP的诸多突变体在酸性环境下很容易发生荧光淬灭,且在很多条件下极易生成有害物质;而DsRed对pH值不敏感,pH值为4.5~12时仍保持稳定,这使其使用范围更加广泛。目前,人们正在研究DsRed对pH不敏感的原因,并将其应用在GFP的优化方面。DsRed更大的缺点是缓慢和不完全的成熟过程及寡聚化。缓慢的成熟过程阻止了它作为报告基因的一些应用,如检测短期的基因表达及生物体快速发育时的一些情况。对GFP进行突变可获得快速成熟的突变株,人们也对DsRed进行类似突变以期得到快速成熟的突变株。

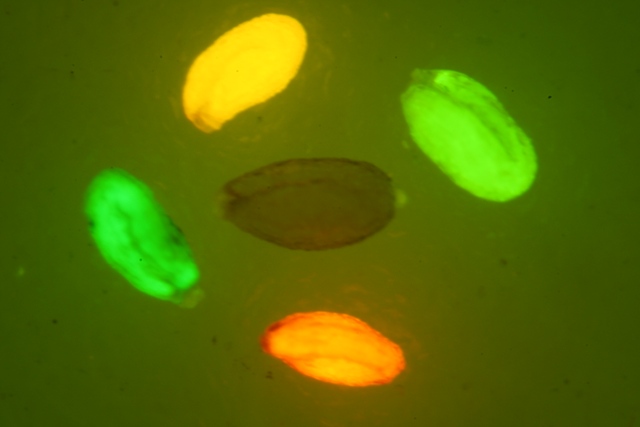
DsRed红色荧光蛋白的激发波长为554nm,发射波长为591nm
基于对DsRed结构的理解和生色机制的阐明,人们采用随机突变或定点突变对DsRed进行优化。与DsRed相比,其突变株的特点主要是减少寡聚化,加快成熟速率和改变波长等。
2.1 减少寡聚化
2.1.1 寡聚化的原因 利用超速离心分析法、能量共振转移法、酵母双杂交法分析DsRed在体内外的表达情况,结果表明,无论在体内还是在体外,DsRed 都倾向形成寡聚体,而且它需要几天时间才能完全成熟。通过时间依赖的荧光各向异性和单分子光谱实验,证明了DsRed即使在纳摩尔级(nm ol?L-1)以下浓度时也存在寡聚体,浓度高时甚至可形成八聚体[4,6]。Yarbrough等[3]和Wall等[7]通过X射线分别得到了DsRed的晶体结构。分析了DsRed的立体结构,发现四聚体表面的疏水基团较少;用计算机分析DsRed四聚体的静电位(势),发现蛋白表面主要带负电荷,而且每个单体的N端均含有几个带正电荷的氨基酸残基,它们通过静电相互作用导致了蛋白的聚集。人们推测每个DsRed四聚体通过4个盐桥与邻近的四聚体相互作用,形成网状的多聚结构。4个配位键的静电相互作用使得“天线样”的结构非常稳定[2]。降低DsRed的寡聚化程度和加快它的成熟过程,可使其具有更好的应用前景,因此,有必要对DsRed进行修饰和改进。
2.1.2 减少寡聚化的方法 人们认为,N端带正电的氨基酸残基可能是引起DsRed聚集的主要原因。因此可通过去除荧光蛋白N端的几个氨基酸残基,或者用带负电荷的或中性氨基酸取代N端的Lys和Arg残基,来获得非聚集的珊瑚虫属(Anthozoa)荧光蛋白的突变体。对DsRed进行定点突变(V105A, I161T,S197A),得到了一个在大肠杆菌中快速及较完全折叠的突变体,命名为E57;然后以E57作母体,进行再次突变。该实验中,三个突变位点(R2A, K5E和K9T)分别以不同的组合进行氨基酸的替换;并用大肠杆菌表达这些突变体,纯化后用聚丙烯酰胺凝胶电泳(PAGE)分析:与母体E57相比,这些突变体的聚集水平均有所下降,R2A突变似乎对聚集水平的下降影响更大。突变株E572NA(R2A,K5E和K9T)的体内外实验表明,该蛋白聚集程度低,而且成熟速率是最快的;它的更大激发波长、荧光强度与母体E57相似。动物实验表明,该蛋白的细胞毒性明显低于DsRed和E57。现在E572NA已经被Clon2 tech公司商业化,名称为DsRed2[2]。而Campbell 等[8]替换了DsRed序列中的33个氨基酸残基,得到了单体红色荧光蛋白mRFP1,该蛋白比DsRed更适于与G FP等进行多色荧光标记,但是,它仍具有荧光强度偏低等缺点,需进一步改进。
2.2 改变发射波长的突变
2.2.1 发射绿色荧光突变株
人们曾对DsRed进行随机和定点突变,得到突变株P37S,K83R,N42H 和T217S,可将DsRed稳定在绿色中间体,更大吸收波长为480nm;另一个中间体的更大吸收波长为408nm,实验证明它们是生色团的质子化和脱质子化形式,这与G FP的生色团类似[4,9]。这些绿色荧光突变株可作为红色荧光生色团形成机制的证据。
2.2.2 发射波长红移(red2shift,RS)突变株 据报道,荧光蛋白发射波长越长,其检测灵敏度越高。人们对mRFP1进行定点突变(突变位点:T202I,R3K, S112T,P188Q,M182K和V96I),得到增强型的单体mRFPmars,其更大吸收和发射波长分别为585和602 nm。而Fradk ov等对两个红色荧光蛋白(dsFP593和drFP583)进行随机突变,得到突变株Ds/drFP616,其发射波长明显延长(616nm),但聚集性还很高,随后对Ds/drFP616进一步突变(K5E和K9T),其突变株Ds/drFP6162K5E/K9T的聚集性明显降低。
2.2.3 发射波长可变的荧光蛋白 最近,人们还发现了一个有趣的DsRed突变体,命名为E5,又称“荧光计时器(fluorescent timer)”。它发射的荧光颜色随蛋白浓度的变化而改变,从鲜绿色、黄色、橙色,最后变成红色[12]。将这种蛋白的基因插入到几种不同类型的细胞基因组中,使它处于某个启动子控制之下,观察基因开启和转录等,从而使研究者方便、快速地了解到基因转录的适时信息。对DsRed2T imer 进行突变(R2A,K5E,K9T),突变体命名为T imer2 NA。它与母体DsRed2T imer的成熟过程及颜色变化相同,实验证明它不形成聚集体[2]。
2.3 适合特定细胞的修饰
Sorensen等[13]利用随机突变的方法,得到可以在细菌中高度表达的快速成熟的DsRed.T3突变株DsRed.T32S4T。用流式细胞仪检测,DsRed.T32S4T 的发光强度足够检测到沙门菌基因的表达;然而,在利什曼原虫和HeLa细胞中,看不到荧光。说明突变引起的mRNA折叠变化影响了原核的转录系统,但对真核转录系统不起作用。而将DsRed基因置于不同生物专用的启动子下,则可以检测该生物的转化效率及基因表达的效率。如将DsRed启动子置于丝状真菌常用的启动子Pgpd之下,得到了载体Pgpd2DsRed,用其转化镰刀菌,可观察到镰刀菌对西红柿的侵染、分布及致病力,还可以用它与含GFP的载体共转化青霉属和木霉属的真菌,得到双标记的真菌。
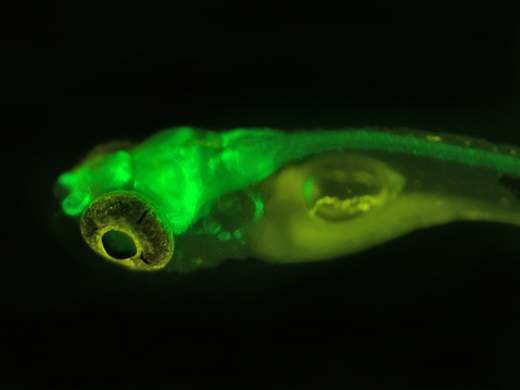
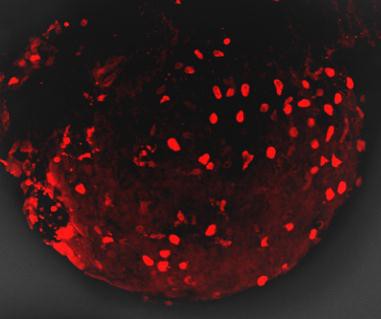
目前,DsRed已应用于启动子分析、转基因、蛋白质和细胞定位、药物筛选、细胞、整个器官的双标记和三标记培养,以及利用荧光共振能量转移(FRET)研究蛋白与蛋白的相互作用;另外,DsRed还可作为胞内不同信使的荧光传感器。为研究ABC转运基因如何在线虫中起作用,人们设计了融合有荧光蛋白的启动子,以观察这些基因何时在体内开启[15];还可将DsRed与蛋白质融合,分析蛋白的表达与定位;或者构建融合有DsRed 的基因组文库或cDNA文库。最近,人们利用转基因技术获得了一个全身发射绿色荧光的小鼠,在小鼠体内移植发射红色荧光的胰腺肿瘤,可清楚地观察到肿瘤和基质的相互作用,这为人们研究肿瘤的发生、发展提供了很好的手段。DsRed与显微镜技术相结合的荧光成像技术,可应用于药物代谢、药剂学等学科,用来分析药物递送载体在组织器官、细胞中的分布和定位等。
4 结语
自1999年红色荧光被发现后,引起了科学家广泛的兴趣,人们对其理化性质和发光机制进行了深入的研究;通过对其修饰,人们得到了聚集程度低、成熟速率更快、发光强度更强和发射波长更长的突变株,这些都极大地方便了DsRed在科学研究中的应用。目前,DsRed的使用还远未有G FP广泛,相信随着对DsRed研究的深入,DsRed将成为人们进行科学研究的重要手段。
如果要观察Dsred红色荧光蛋白(red Fluorescent Protein)的表达,美国路阳生产的便携式荧光蛋白激发光源可以选择LUYOR-3260GR和LUYOR-3415(X)G系列双波长荧光蛋白激发光源。红色荧光蛋白采用绿光激发,佩戴LUV-50A红色观察眼镜观察,如希望提供更多详细信息,可直接联系上海路阳生物技术有限公司的销售客服。
 关注我们
关注我们