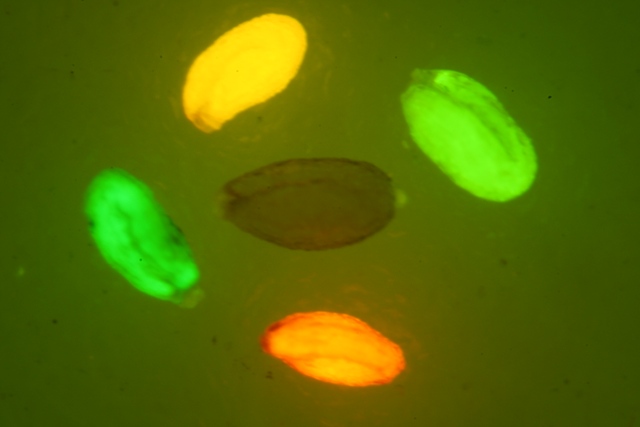
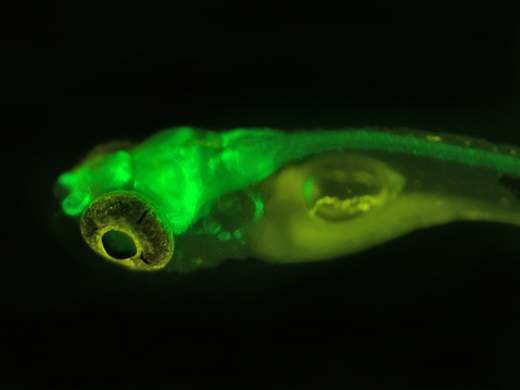
LUYOR-3109高强度紫外催化光源促销
LUYOR-3109紫外光源采用了9颗365nm大功率led,安装有二次光学透镜,输出紫外线强度高,...
2024-08-08

作者:激发光源事业部时间:2019-12-08 11:32:28浏览7442 次
天然gfp的荧光强度低,表达易受温度影响,通过改变发色团氨基酸种类和排列顺序,获得了改良型的gfp。将第65位的ser突变成thr,可显著增加荧光强度和稳定性;第66位氨基酸tyr突变为his或trp,荧光波长变短,可分别得到发蓝光和蓝绿光的gfp;改变发色团周边氨基酸的种类,也可以使荧光波长发生变化,将第203位的thr突变成tyr,就可获得黄色荧光。
绿色荧光蛋白在植物病理学研究中的应用
报告基因(reportergene)是编码某种检测蛋白或酶的基因,通过它的表达产物来标定目的基因的表达调控情况,具有灵敏度高、可信度好和检测方便。报告基因种类繁多,如氯霉素乙酰转移酶(chloramphenicolacetyltransferase,cat)基因、分泌型碱性磷酸酶(secretedalkalinephosphatase,seap)基因、绿色荧光蛋白(green fluorescentprotein,gfp)基因、β-葡萄糖苷酶(β-glucosidase,gus)基因、β-半乳糖苷酶(β-galactosidase,β-gal)基因和萤火虫荧光素酶(fireflyluciferase,fl)基因等。由于gfp具有荧光性质稳定、观察方便、对细胞无毒害、无物种特异性、不受假阳性干扰和无需底
物等优点,深受科研和检测人员的青睐,目前已成为分子生物学领域的重要标记蛋白。
绿色荧光蛋白于1962年在维多利亚多管发光水母(aequoreavictoria)中发现,该蛋白在紫外光照射下发出明亮的绿光;1992年prasher成功克隆gfp的全长cdna,2年后将gfp基因作为报告基因导入大肠杆菌(escherichiacoli)和秀丽隐杆线虫(caenorhabditiselegans)中表达,发现在无任何辅助底物的情况下,gfp可以在活细胞中发光;通过基因突变和化学修饰对gfp基因进行改造,解决了gfp荧光强度低、光选择性差和光谱狭窄等缺陷,目前荧光颜色已涵盖紫色到红色的所有可见光区域。近年来,gfp基因在启动子分析、转基因监测、药物检定、污染物示踪、抗病检测和基因表达调控等方面得到了广泛应用。gfp基因在植物病理学研究中也得到了广泛应用,本文将展开相关论述。

gfp的发现
生物发光是一种普遍现象,自然界中,大约有700种生物能够发光,包括细菌、大型真菌和动物,约80%的发光生物生活在海洋中,它们通过发光来寻找食物、逃避敌害和吸引异性。生物发光通常需要底物的参与,如萤火虫发光是通过荧光酶)催化荧光素(luciferin)来实现的,而gfp是一种能在蓝色光线激发下发出绿色荧光的特殊蛋白质,在发现初期并未受到重视,它只是研究水母素(aequorin)时的副产品。20世纪90年代,gfp在大肠杆菌中成功表达,开创了该蛋白的应用先河,之后在生命科学多个领域中得到了广泛应用,并取得了令人瞩目的成就。
gfp的结构与发光原理
野生型gfp基因组全长为2600bp,由3个外显子和2个内含子组成,编码区全长为717bp,分别以atg和taa为起始和终止密码子,编码238个氨基酸,分子量为27ku。gfp的三维结构呈圆筒形,11个β折叠围在四周,1个α螺旋贯穿整个圆筒结构,其中3个氨基酸残基ser65-tyr66-gly67构成发色团,位于圆筒中央并附着在α螺旋上[12]。尽管在氨基酸水平上仅23%的相似性,但gfp的结构与从discosomasp.分离到的红色荧光蛋白(redfluo-rescentprotein,rfp)十分相似,后者也是由11个β折叠和1个α螺旋组成,形状也呈折叠筒状[12]。gfp的发光原理是:gly67的酰基和ser65的羧基经亲核反应生成咪唑基,tyr66通过脱氢使芳香团与咪唑基结合,形成对羟基苯甲酸咪唑环酮生色团发出荧光。
天然gfp的荧光强度低,表达易受温度影响,通过改变发色团氨基酸种类和排列顺序,获得了改良型的gfp。将第65位的ser突变成thr,可显著增加荧光强度和稳定性;第66位氨基酸tyr突变为his或trp,荧光波长变短,可分别得到发蓝光和蓝绿光的gfp;改变发色团周边氨基酸的种类,也可以使荧光波长发生变化,将第203位的thr突变成tyr,就可获得黄色荧光。
gfp在植物病理学研究中的应用
随着成像技术、检测手段的快速发展,gfp基因及其突变体作为报告基因在分子生物学领域中得到了大规模应用。gfp基因在植物病理学方面主要体现在抗病基因的亚细胞定位、启动子活性分析、病原菌-植物互作研究和基因表达分析等方面。
3.1抗病基因的亚细胞定位
真核细胞具有复杂亚细胞结构,蛋白质在细胞内的定位是细胞生物学研究的热点之一。亚细胞定位能明确表达蛋白或产物在细胞内的出现部位,常用的方法有免疫荧光技术、胶体金标记免疫电镜技术和gfp融合蛋白技术等。免疫荧光技术将免疫学方法与荧光标记技术相结合研究特异蛋白抗原在细胞内分布,通过检测荧光素发出的荧光对组织或细胞中的抗原进行细胞定位,这种方法具有简便、快速和特异等优点,缺点是敏感性低、定量困难和易受化学发光物质干扰[16]。胶体金标记免疫电镜技术是以胶体金作为示踪标志物或显色剂,利用其在碱性条件下带负电的特点进行抗体标记,具有特异性强、灵敏度高和定位正确等优点,缺点是实验步骤多、技术难度大。而gfp具有荧光性质稳定、无物种特异性、对细胞无毒、灵敏度高和能在细胞中稳定表达等优点,gfp融合蛋白技术已成为亚细胞定位的重要手段。
npr1基因是拟南芥(arabidopsisthaliana)系统获得抗性中的一个关键基因,它在植物抗病反应中起到重要作用,国内外许多学者对其进行了较系统的研究,利用农杆菌介导法将npr1-gfp融合基因导入拟南芥,结果表明,npr1基因在细胞核中表达,在病菌诱导下,npr1-gfp整合蛋白的积累量增加[17]。osWrky13是水稻(oryzasativa)的一个转录因子,过量表达能提高对白叶枯病和稻瘟病的抗性,将gfp-osWrky13融合基因导入洋葱表皮,结果表明该基因在细胞核中表达,并进一步证明osWrky13具有dna结合活性[18]。过氧化物酶在抗病反应中起着重要的调控作用,将一粒小麦(triticummonococcum)过氧化物酶基因tmprX8与gfp融合后导入洋葱表皮,融合蛋白出现在液泡中。
3.2启动子活性分析
启动子是一段直接与rna聚合酶及转录因子相结合,决定基因转录起始与否的特定dna序列,采用有效的启动子能够显著提高外源基因在植物中的表达水平,因此启动子活性检测是植物基因工程的重要内容之一。传统的启动子活性检测采用β-葡萄糖苷酸酶基因、氯霉素乙酰转移酶基因和冠缨碱合成酶基因等作为报告基因,检测时需要特定的底物和辅助因子,而且对植物组织或细胞具有破坏性。gfp是启动子分析的新型报告基因,利用荧光显微镜观察基因表达情况,能较正确地鉴定启动子活性大小,并达到量化的目的。
苏云金芽胞杆菌(Bacillusthuringiensis)是一种应用广泛的杀虫微生物,已成功地用于防治农业、林业、贮藏物害虫及医学昆虫,随着微生物农药需求量的增大和人类环境保护意识的增强,生物安全日益受到人们的重视。利用gfp基因研究了苏云金芽胞杆菌pcry3a和pBtⅠ_BtⅡ等启动子的活性,为有效选择驱动标记基因更优的表达体系和进一步构建、安全的基因工程菌创造了条件[21]。利用gfp基因作为报告基因,采用基因枪法进行洋葱表皮遗传转化,培养24h后检测基因表达情况和荧光强度,能正确测定出启动子活性的大小。
3.3病原菌与植物互作研究
病原菌侵染植物过程中,病原菌无毒基因产物与寄主植物抗病基因产物相互识别和结合,产生的信号分子通过传导,激活一系列抗病基因的表达,引发复杂的生理生化反应,从而表现出抗病特性[22]。深入研究植物与病原菌的识别方式、亲和性或非亲和性的互作模式,对揭示植物-病原菌互作机制的认识具有重要意义[23]。研究病原菌与植物互作的传统方法有组织印迹法、放射性标记核酸探针法和gus染色法等,这些均为非活体检测,因此有一定的局限性[21]。而gfp能有效克服这一弊端,可直观、实时监测病原菌的发生、定殖和侵染过程,目前已成为研究病原菌与植物互作的有力工具[25]。烟草黑胫病是烟草生产过程中的重要病害,世界各地均有发生,利用原生质体介导法将gfp基因导入病原菌(phytophthoraimperfectavar.nicoti-anae)中,观察到了烟草黑胫病菌的生长动态和与寄主互作方式,这是gfp作为报告基因在病原微生物研究中的应用。香蕉叶斑病和斑枯病由球腔菌属(Mycosphaerella)真菌引起,其中香蕉黑条叶斑病菌(M.fijiensis)和香蕉黄条叶斑病菌(M.musi-cola)为叶斑病的病原菌,而M.eumusae为斑枯病的病原菌,将gfp基因导入这3种真菌,清楚地观察到了病菌在叶片组织中的生长动态,并且发现随着菌丝的侵入叶片逐渐褪绿和坏死。草莓炭疽病是草莓(fragariaananassa)生产的重要病害,近年来发生十分严重,尖孢炭疽菌(colletotrichumacutatum)是草莓炭疽病的病原菌,它侵染叶片、匍匐茎、花蕾、萼片和果实,用电穿孔法将gfp基因导入萌发的分生孢子研究草莓致病炭疽病原菌和非致病炭疽病病原菌的生活规律,为阐明致病机理奠定了基础。
利用含gfp基因和氯霉素抗性的质粒分别转化对黄瓜枯萎病菌(fusariumoxysporum)有明显抑制作用的生防菌株BrevibacillusbeviszJy1和BacillussubtiliszJy-116,得到具有gfp绿色荧光和氯霉素抗性的转化子,成功检测了菌株在田间定殖的可行性,为菌株的实际开发利用提供了依据。以莴苣盘梗霉(Bremialactucae)hsp70的启动子和ham34基因的终止子构建gfp表达载体,利用原生质体介导法导入重组载体,观察到gfp在不同时期大豆疫霉(phytophthorasojae)中的表达情况,同时明确了大豆疫霉在大豆叶片、下胚轴和根部的侵染行为新霉素磷酸转移酶基因(hpt)、潮霉素磷酸转移酶基因(npt)、链霉素磷酸转移酶基因(nptii)、草铵膦(glufosinate)抗性基因(bar)、草甘膦(glyphosate)抗性基因(epsps)等为传统的转基因植株选择标记[32],通常需要6~8周或更长的时间才能鉴定出潜在的转基因苗,并经southern杂交分析来确定是否为转基因阳性植株,整个过程花费时间较长。使用抗生素、除草基因作为转基因标记是否会引发生物安全问题是人们普遍关注的问题,而gfp基因是一种安全、可靠、省时的标记基因,不会产生“超级杂草”和“基因逃逸”等生态问题,可替代抗素、除草剂快速筛选转基因植株或组织,还可通过标记花粉来研究转基因植物花粉漂移和授粉过程。
sgt1(suppressoroftheg2alleleofskp1)是植物抗病信号传导途径中的重要调控基因,参与多种植物抗病基因介导的抗病信号途径,sgt1基因的突变或沉默会导致多种植物r基因介导的抗病性的丧失,sgt1还参与调控植物的非宿主抗性[34-35]。以gfp为报告基因对atsgt1a和ats-gt1b的功能进行了研究,通过荧光检测,明确了这两个抗病基因的表达部位和表达量[36]。利用peg法将来自水稻的抗病基因Xa21导入柑橘原生质体,通过gfp荧光检测获得愈伤组织和抗性芽,经微嫁接后获得了抗病植株[37]。稻黄单胞菌(Xan-thomonasoryzaepv.oryzae)是水稻白叶枯病的病原,以gfp为报告基因,利用农杆菌介导法将来自水稻的抗病基因Xa27导入,结果表明Xa27对水稻黄单胞菌具有抗性作用[38]。
展望
在对gfp进行突变改造后,现已获得发蓝光的Bfp、发黄光的yfp和发青色光的cfp,研究人员可根据需要进行选择[4]。值得一提的是,1999年从珊瑚虫(discosomagenu)中分离到了与gfp同源的蛋白质dsred,它在紫外线照射下发出红色荧光,尽管dsred与gfp在氨基酸水平上仅23%的相似度,但三级结构十分相似[12]。与gfp相比,dsred的激发和发射波长较长,发射峰位于培养基、组织培养器材及细胞成分等产生的荧光背景范围之外,具有较高的信噪比,而且在细胞内荧光转换效率高,检测更为容易[39]。随着研究的进一步深入,一定能开发出更明亮、更稳定、穿透性更强的荧光蛋白,它们将成为生命科学研究的重要工具。
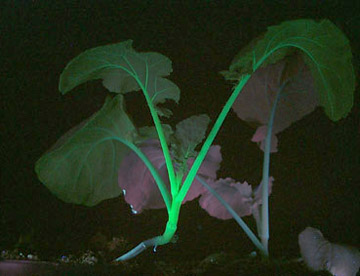
实验室从青花菜(Brassicaoleraceavar.itali-ca)中克隆到了多个霜霉病(hyaloperonosporaparasitica)抗性相关基因(downymildewresist-ance-relatedgene,dMr),并以gfp为报告基因,对其中的BodMr1进行了遗传转化研究。在构建重组载体pBi121-gfp-BodMr1和将其导入农杆菌lBa4404的基础上,经共养、分化培养、生根培养、荧光检测和炼苗,成功获得了17棵转基因青花菜阳性植株,这些植株在紫外线照射下发出明亮的绿色荧光,抗病性检测结果证实这些植株对霜霉菌的抗性均有所增加,个别达到高抗水平(待发表)。gfp的引,为快速、有效筛选青花菜转基因阳性后代及选育抗病材料提供了保障。
 关注我们
关注我们