LUYOR-3109高强度紫外催化光源促销
LUYOR-3109紫外光源采用了9颗365nm大功率led,安装有二次光学透镜,输出紫外线强度高,...
2024-08-08

作者:时间:2019-11-23 20:42:26浏览4935 次
外源基因导入宿主细胞后有两种表达模式, 一是整合到宿主细胞基因组中稳定表达; 二是以质粒形式导入宿主细胞后瞬时表达。由于后者快速且方便已被广泛应用到基因工程中)。植物瞬时表达系统不会产生基因沉默、基因漂移等, 可有效避免基因污染, 是检测外源基因表达情况的理想方法。棉花外源基因的稳定表达是个复杂的长期过程,建立一个快速、经济、安全的瞬时表达系统对研究棉花基因功能有重要意义。
植物瞬时表达系统在研究功能基因的亚细胞定位、蛋白质互作、启动子活性等方面具有重要作用。选取棉花(Gossypium hirsutum)子叶为材料, 以β-葡糖醛酸酶基因(GUS)、增强型绿色荧光蛋白基因(eGFP)为报告基因, 对棉花子叶生长时间、农杆菌浓度、注射量、共培养时间等因素进行了分析, 建立了农杆菌介导的棉花子叶瞬时表达系统。结果表明,对萌发6天的棉花子叶注射50 μLOD600为0.5的含目的基因的农杆菌LBA4404或GV3101, 共培养2–3天后, 外源基因表达效率更高。利用此方法从棉花萌发到观测结果, 仅需8–13天, 且操作简单易行, 可快速检测棉花来源的启动子活性、亚细胞定位、蛋白质互作等。该方法在棉花基因功能研究方面有很好的应用价值。
棉花(Gossypium hirsutum)是世界纺织业的主要原材料, 有很高的经济价值和应用价值。近年来, 棉花的基因组测序已经完成, 其中有44.9 Mb的基因已被注释(Paterson et al., 2012)。随着注释基因的不断增多, 需要寻找一种快速有效的方法对基因功能进行分析。使用瞬时表达系统操作简单, 无需特殊仪器设备, 外源基因的表达效率高且安全, 不易产生基因漂移(邱初等, 2009; Grefen et al., 2010; 蓝岚等,2013)。同时此方法可有效避免基因沉默等, 在基因功能分析及亚细胞定位方面应用广泛(Grebenok etal., 1997; Li et al., 2011)。
目前, 农杆菌介导的植物瞬时表达系统已在许多植物中建立并应用, 如大豆(Glycine max)、拟南芥(Arabidopsis thaliana)(Koroleva et al., 2005)、红花(Flos carthami)(Belide et al., 2011; 杨晶等, 2011)、油菜(Brassica campestris)(谭小力等, 2012)以及玉米(Zea mays)、水稻(Oryza sativa)等(李浩戈等,2012)。近年来, 棉花中瞬时表达系统主要利用愈伤组织经酶解处理后形成的原生质体来完成。该方法能快速检测棉花功能基因的瞬时表达、亚细胞定位等,但操作繁琐, 需要基因枪进行外源基因的转化, 实验成本高(李杰等, 2003; Shibasaki et al., 2012)。本实验以棉花子叶为材料, 利用农杆菌注射法建立了瞬时表达系统, 确定了子叶的更佳生长时期、注射菌量以及共培养时间等。在此基础上, 构建了35S-eGFP(pG3)表达载体, 对棉花子叶进行了转化。实验结果表明, 该方法不仅适用于带GUS标签的基因, 也适合带eGFP标签的基因功能研究。上述方法的建立有助于棉花功能基因的亚细胞定位、蛋白质互作以及启动子活性等研究。
1 材料与方法
1.1 实验材料
棉种柯字312(Gossypium hirsutum cv. ‘Coker 312’)系河北农业大学马峙英教授馈赠。pGEM-T Easy载体购自Promega 公司。农杆菌(Agrobacterium tumefacien)LBA4404菌株由北京大学朱玉贤教授惠赠。含pCAMBIA2301的GV3101菌种由江苏大学生命科学学院谭小力教授惠赠。含eGFP的pUCegfpC质粒为西北农林科技大学王倩老师惠赠。农杆菌GV3101菌株、大肠杆菌(Escherichia coli)DH5α菌株为本实验室保存。
1.2 实验方法
1.2.1 植物培养
选取饱满的种子, 在37°C水浴中浸泡14小时, 播种于盛有培养土(蛭石:营养土=1:3, v/v)的小钵中, 覆膜后于28°C、光暗周期为16小时/8小时的室内培养。待发芽后去膜, 生长数天后用于实验。
1.2.2 子叶注射及共培养
吸取200 μL含pCAMBIA2301的GV3101菌液添至10mL含50 mg·L–1卡那霉素(Kan)、50 mg·L–1利福平(Rif)和20 mg·L–1庆大霉素(Gent)的LB液体培养基中,于28°C、每分钟180转振荡培养至所需OD600值时,10 000 ×g离心1分钟。收菌, 除去上清, 用含100μmol·L–1乙酰丁香酮与10 mmol·L–1硫酸镁的MS侵染液悬浮菌体, 得到转化液。挑选生长状态良好且生长一致的棉花子叶, 用蒸馏水清洁叶片背面, 蘸75%乙醇擦拭, 再用蒸馏水清洗残留乙醇。选用背面叶脉部位, 用去掉针头的注射器注射转化液, 注射完毕后用蘸有蒸馏水的棉签清洁叶片背面。将注射后的棉株放入25°C、光暗周期为16小时/8小时的培养箱中培养。
1.2.3 GUS染色及活性检测
将共培养一定天数的子叶用蒸馏水清洗后置于GUS染色液中( 每100 mLGUS 染液中含有2.1 mmolNaH2PO4, 2.9 mmol Na2HPO4, 0.5 mmol EDTA,pH7.0; 0.1 mmol KFe(CN)4, 0.1 mmol KFe(CN)3,0.1 mLTritonX-100, 0.1 g X-Gluc), 于37°C每分钟180转振荡染色24小时, 然后置于真空泵中, 0.08MPa抽真空30分钟, 继续振荡染色48分钟, 除去染色液。将子叶置于无水乙醇中脱色8小时, 观察并拍照。
用MUG法(Viana et al., 2011)提取不同处理后且共培养一定天数的棉花子叶总蛋白。吸取上清100μL, 加入到400 μL经37°C预热的GUS提取缓冲液(每100 mLGUS提取缓冲液中含有50 mL 0.1 mol·L–1 pH7.0的磷酸缓冲液, 1 mL 10% SDS, 2 mL 0.5 mol·L–1pH 8.0的EDTA, 100 μLTriton X-100, 100 μLβ-巯基乙醇), 再加入500 μL MUG底物(8.8 mg MUG, 溶于10 mLGUS酶提取液中), 37°C温浴。立即取出200 μL加入到800 μL反应终止液(0.2 mol·L–1Na2CO3)中混匀, 此标记为1号管, 在室温下避光保存。此为反应0时的样品, 荧光测定时以此为空白, 并开始计时。将反应管放入37°C水浴中进行酶反应, 60分钟后, 取200 μL反应液, 加入到800 μL反应终止液中混匀, 此标记为2号管, 为反应60分钟时的样品。用多功能微孔测量仪(美国EnSpire)在激发波长365 nm、发射波长455 nm、狭缝10 nm时测定不同时间点的荧光强度值。根据4-甲基伞形酮(4-MU)制作的标准曲线得出GUS活性(Y=(C2号管–C1号管)/C总蛋白/时间(h))。
1.2.4 35S-eGFP融合表达载体的构建
以pUCegfpC质粒为模板, 上、下游引物分别为eGFP1_F(5′-TCTGAATTCATGGTGAGCAAGGGCGAG-3′)和eGFP2_R(5′-TTCTCTAGATTACTTGTACAGCTCGTCC-3′) 。用高保真酶PrimeSTAR(Ta-
KaRa)扩增获得eGFP基因序列(5′端和3′端分别含EcoRI和XbaI酶切位点), 连入pGEM-T Easy载体。测序验证后将该质粒命名为pG1 。将质粒pG1 与pKannibal分别用EcoRI/XbaI酶切, 回收相应片段连接并测序后得到pG2质粒。将pG2用NotI酶切, 回收35S-eGFP片段, 连接到pART27载体中, 转化大肠杆菌DH5α, 经蓝白斑筛选获得阳性克隆, 测序正确后命名为pG3。采用冻融法将pG3转入农杆菌LBA-4404和GV3101中, 并分别命名为LBA-pG3和GVpG3。
1.2.5 农杆菌转化及eGFP荧光检测
将农杆菌LBA-pG3和GV-pG3培养至OD600为0.5, 收集3 mL菌液加入900 mL MS培养基和100 mL乙酰丁香酮悬浮后使用。挑选生长6天的棉花幼苗, 在其子叶背面注射含LBA-pG3或GV-pG3的农杆菌转化液。棉花子叶共培养1–11天后, 采集叶片, 置于倒置荧光显微镜(莱卡DMIL LED)下, 在490 nm蓝色激发光下观察eGFP的表达。
1.2.6 半定量PCR检测
采集共培养3天的棉花子叶, 用打孔器在注射位置取材, 保证材料直径均为1.3 cm。用试剂盒(Plant RNAKit, OMEGA Bio-tek)提取总RNA。取1 μg RNA用逆转录试剂盒(Prime ScriptTM RT Reagent Kit PerfectReal Time)逆转成cDNA。以cDNA为模板, 棉花Actin为内参基因( 上、下游引物分别为Actin_F, 5′-AGCTATGAGTTGCCTGATGG-3′; Actin_R, 5′-ATTGTAAGTGGTCTCGTGAATA-3′), 以eGFP_F(5′-AGAACGGCATCAAGGTGAAC-3′) 和eGFP_R1(5′-TGCTCAGGTAGTGGTTGTCG-3′)分别作为上、下游引物进行PCR扩增。PCR反应体系(总体积为25μL): 12.5 μL 2×Taq PCR Mix, 1 μL cDNA, 上、下游引物(10 μmol·L–1)各2 μL, 7.5 μLddH2O。PCR反应程序为: 95°C预变性5分钟; 95°C变性1分钟, 60°C复性1分钟, 72°C延伸1.5分钟, 30个循环; 72°C终延伸10分钟。反应产物于4°C保存。
1.2.7 免疫组织化学检测eGFP蛋白
采集共培养3天的棉花子叶, 用打孔器取直径为1.3cm的组织圆片, 经甲醇脱色、崩溃酶(Sigma)裂解细胞壁以及0.08 MPa抽真空30分钟, 增加叶片渗透性后, 加入eGFP抗体, 37°C孵育3小时, 漂洗后加入显色剂DAB, 30分钟后用蒸馏水洗涤以停止反应。具体步骤参见文献(Sauer et al., 2006)所述。
2 结果与讨论
2.1 棉花子叶瞬时表达系统的建立
不同生长时期的子叶对农杆菌的侵染和外源基因的表达有重要的影响。本实验采用萌发后生长2、6、10、14 和18 天的子叶为材料, 用含pCAMBIA2301 的GV3101菌液(50 μL农杆菌, OD600=0.5)(简称含GUS基因的农杆菌)注射棉花子叶的背面叶脉部位, 共培养2天后, 取整片子叶进行GUS染色。结果(图1A)显示, 随着子叶生长期的延长, GUS染色逐步明显, 至
第6天达到高峰, 之后GUS染色呈现降低趋势, 至18天时几乎不显色。其中, 注射部位及其周围(0.2–1.0cm)为GUS显色部位。同步采集注射后共培养2天的子叶, 用MUG法定量分析GUS酶活性, 结果与上述定性分析结果具一致性(图1B)。实验表明生长6天的子叶最适合转化。

不同生长天数的棉花子叶GUS染色结果(A)及定量分析结果(B)
有文献报道农杆菌浓度对叶片侵染的结果有直接影响(张福丽等, 2012)。本实验选取OD600为0.3、0.4、0.5和0.6的含GUS基因的农杆菌50 μL分别对萌发后生长6天的子叶进行注射, 共培养2天后, 进行GUS染色(具体操作同上)。结果(图2A)显示, 随着注射菌液浓度的增加, GUS染色的色斑、变得明显, 至注射浓度OD600为0.5时达到更佳状态, 之后GUS染色呈现下降趋势。同步采集注射后共培养2天的子叶,提取总蛋白, 定量分析其中GUS蛋白含量, 结果与上述定性分析结果一致(图2B)。实验表明注射浓度OD600为0.5时染色最明显, 此时的转化效率更高。为了确定菌液更佳注射量, 用OD600为0.5的含GUS基因的农杆菌注射液, 对培养6天的棉花子叶进行注射, 注射量分别为20、30、50、100和150 μL。共培养2天后, 进行GUS染色(具体操作同上)。结果(图3A)显示, 随着注射量的逐步增加, 子叶的显色部位有明显变化, 至注射量为50 μL时染色效果更好,注射量为100和150 μL时GUS染色效果下降。同步采集注射后共培养2天的子叶, 定量分析其中的GUS蛋白含量, 结果与上述定性分析结果一致(图3B)。实验表明OD600为0.5的农杆菌注射量为50 μL染色效果更好, 此时的转化效率更高。

本实验还探讨了不同共培养时间对侵染结果的影响。用50 μLOD600为0.5含GUS基因的农杆菌注射液对培养6天的子叶进行注射, 分别共培养1、2、3和4天后, 进行GUS染色(具体操作同上)。结果(图4A)显示, 随着共培养时间的延长, GUS染色逐渐明显。其中在共培养2天时, GUS染色效果更好, 随后染色效果降低。同步采集注射后共培养2天的子叶, 定量分析其中的GUS蛋白含量, 结果与上述定性分析结果一致(图4B)。实验表明共培养2天GUS染色效果更好, 此时的转化效率更高。

2.2 棉花子叶瞬时表达eGFP的荧光检测
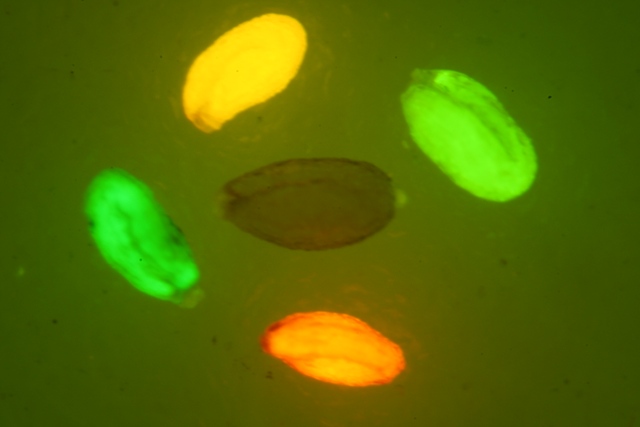
采集注射农杆菌LBA-pG3和GV-pG3后共培养1–11天的棉花子叶, 置于倒置荧光显微镜下观察绿色荧光蛋白的表达。结果(图5)显示, 随着共培养时间的延长,绿色荧光强度呈逐渐增强的趋势, 至3–5天后荧光强度下降。其中注射LBA-pG3的处理组中, 在共培2天时有微量表达, 至3天时观察到的绿色荧光强度最强, 且显著优于同期注射GV-pG3的处理组。实验结果表明eGFP的表达在3天时效果更佳, 且棉花子叶瞬时表达系统选择注射农杆菌LBA4404介导的外源基因表达效果比GV3101更好。

2.3 棉花子叶瞬时表达eGFP的定量分析
为了进一步确定上述实验中所观察到的绿色荧光是eGFP表达的结果, 本实验选取注射后共培养3天的子叶用半定量PCR法进行RNA水平的检测及免疫组化分析。以cDNA为模板, Actin基因为内参, 对eGFP基因表达进行半定量PCR检测(图6A)。以质粒为阳性对照, 注射不含目的基因的空载农杆菌LBA4404为阴性对照, 结果显示, 在cDNA模板量相同的条件下,阴性对照中扩增不到目的条带, 而注射LBA-pG3能扩增出eGFP条带。表明注射农杆菌LBA4404介导的eGFP能在棉花子叶中表达, 且在共培养3天时表达效果较好。为了进一步确定eGFP蛋白的表达情况,
本实验还进行了棉花叶片免疫组织化学检测, 以显示eGFP蛋白的位置及含量。取共培养3天的棉花子叶,经处理增加叶片渗透性后, 加入eGFP抗体孵育, 用SABC染色试剂盒进行检测, 结果如图6B–D所示。图6中深色圆点为细胞核, 小圆点及染色部分为表达的eGFP蛋白。与对照组(图6B)相比, 生长6天的棉花子叶注射50 μLOD600为0.5的LBA-pG3后, 培养3天, 确有eGFP蛋白的表达。其中, 农杆菌注射部位1.3 cm区域内的eGFP蛋白的表达量(图6D)大于注射部位1.3 cm区域外eGFP蛋白的表达量(图6C)。上述结果表明棉花瞬时转化系统可以成功表达外源基因。

2.4 讨论
外源基因导入宿主细胞后有两种表达模式, 一是整合到宿主细胞基因组中稳定表达; 二是以质粒形式导入宿主细胞后瞬时表达。由于后者快速且方便已被广泛应用到基因工程中(Shibasaki et al., 2012)。植物瞬时表达系统不会产生基因沉默、基因漂移等, 可有效避免基因污染, 是检测外源基因表达情况的理想方法。棉花外源基因的稳定表达是个复杂的长期过程(Chakravarthyet al., 2014 ), 建立一个快速、经济、安全的瞬时表达系统对研究棉花基因功能有重要意义。
刘雪梅, 王蕾, 文添龙, 何鹏, 俞嘉宁 (2014). 农杆菌介导的棉花子叶瞬时表达系统的建立. 植物学报 49, 587–594.
 关注我们
关注我们